Dyslexia treatment program based on four pillars: (1.) the Orton-Gillingham approach, (2.) cognitive training, (3.) the development of two crucial brain areas for reading, and (4.) solid learning principles.
Table of contents:
- Introduction
- A dyslexia treatment program based on four pillars
- Orton-Gillingham as a treatment for dyslexia
- Dyslexia treatment and cognitive training
- Dyslexia treatment targeting two brain regions
- Dyslexia treatment and learning principles
- The bottom line
- Key takeaways
Introduction

Why many intelligent children should suffer seeming “word blindness” has baffled and frustrated parents and professionals in many fields for more than a century.
“Word blindness” was the first term used to describe a reading disability. Dyslexia, which is the term we use today, is a difficulty with words. Like all words that start with dys it refers to a problem, deficiency, or lack. Lexia has to do with words and language. Dyslexia thus refers to difficulty with reading and writing words. Children with dyslexia cannot read without stumbling, their spelling may be atrocious, and they are often innumerate too — sometimes after years of tutoring and treatment.
Edublox’s dyslexia treatment program differs from other dyslexia interventions as it is based on four pillars. Is there, at last, some light at the end of the tunnel?
Dyslexia treatment program based on four pillars
One nigth ther saw a firer in a builing one man saw very injure he was rushed to hositipal and theree day he diad for interle njres.
(One night there was a fire in a building. One man was very (badly) injured. He was rushed to hospital and (after) three days, he died from internal injuries.
The above transcript is not a misprint. You are reading a sentence written by a person with dyslexia — complete with the more common characteristics like lack of punctuation, misspelling, reversal of words and letters, mixed-up sentence structure, and poor grammatical construction. The sentence is practically incomprehensible to the average reader, and the partially corrected and punctuated translation below is necessary for interpretation.
In our age of verbal and written communication, linguistic and reading difficulties are not easy to cope with. It’s a problem that can leave one in a lonely world, feeling unconfident and insecure. Children’s self-esteem is often bludgeoned out of existence when mocked by their peers and dubbed lazy, stupid, or careless by exasperated teachers or parents.
When, in the 1960s, the term ‘dyslexia’ came to be widely used to describe typical children with learning difficulties akin to those who had suffered brain damage, it was seized on with relief by parents and teachers alike. The idea that there was a physical syndrome called dyslexia, and that it was only a matter of time before its cause could be isolated and a solution developed, seemed soothingly feasible.
Multiple specialists have produced a sea of research to find the cause and a solution. As a result, there is a labyrinth of differing, opposing, and often contradictory theories about dyslexia — what it is, its causes, and its possible correction. However, despite knowing what cognitive skills are involved when children learn to read and write, and a better understanding of the “reading” brain, dyslexia is still considered an incurable, persistent, and life-long condition (Novembli & Azizah, 2019; Knowles, 2017).
Perhaps the solution to this learning disorder does not lie in one aspect alone but in combining several different approaches, methods, and principles. Edublox’s dyslexia treatment program is based on four pillars: (1.) the Orton-Gillingham approach, (2.) cognitive training, (3.) the development of two crucial brain areas, and (4.) solid learning principles.
1. Orton-Gillingham as a treatment for dyslexia
Edublox’s dyslexia treatment program is aligned with the well-respected Orton-Gillingham approach, developed by Anna Gillingham and founded on the theoretical work of the American neurologist Samuel Orton.

Orton cultivated a particular interest in children with dyslexia. He studied over 1,000 children, and his observations persuaded him that children with dyslexia were especially prone to left-right confusion and reversals, such as mistaking b for d or was for saw. Orton thus concluded that dyslexia was due to a failure to establish a left-right sense, which was, in turn, caused by incomplete cerebral dominance. Gillingham translated these theories into a highly structured reading method.
The Orton-Gillingham (OG) method is a direct, explicit, multisensory, structured, sequential, diagnostic, and prescriptive way to teach reading, spelling, and writing. It is most properly understood and practiced as an approach, not a method, program, or system. In the hands of a well-trained and experienced instructor, it is a powerful tool of exceptional breadth, depth, and flexibility.
The OG approach is one of several reading methods that emphasize the phonetic regularities of English in its instructional sequence. Initially, individual letters are taught using drill cards, whereafter, students are taught to blend them into simple consonant-vowel-consonant words (e.g., map, hit, Tim). The similarly structured introduction of subsequent single letters, blends (e.g., st, cl, tr), and other letter combinations (e.g., sh, ea, tion) are meticulously sequenced. Later stages in the sequence include sentence and story writing, syllabication, dictionary skills, and advanced spelling rules.
The outcomes not statistically significant
Ritchey and Goeke (2006) reviewed 12 studies that included elementary students, adolescents, and college students.
Five of the 12 studies reported that the OG instruction was more effective than the control interventions for all measured outcomes. Four reported that the OG instruction was more effective for at least one (but not all) outcome than other interventions. Two reported that the alternate instruction was more effective than the OG instruction and found no significant differences once variables were included.
The largest effects were reported for word attack or nonword reading outcomes, with a mean effect size of .82, and comprehension outcomes, with a mean effect size of .76. Other mean effect sizes were in the small to medium range.
Stevens and colleagues (2021) conducted a meta-analysis to examine the effects of OG reading interventions on the reading outcomes of students with or at risk for word-level reading disabilities (WLRD). The purpose of a meta-analysis is to systematically combine and analyze data from published research studies to understand what that body of research says about a particular question.
The findings of Stevens et al.’s meta-analysis suggest that OG reading interventions do not statistically significantly improve foundational skill outcomes (i.e., phonological awareness, phonics, fluency, spelling). However, the mean effect size was in favor of OG-based approaches. Similarly, there were no significant differences in vocabulary and comprehension outcomes for students with or at risk for WLRD.
Solari et al. (2021) summarize the results of the studies mentioned above:
The evidence in favor of OG programs is limited, either due to a mix of positive and negative effects or, more frequently, because available studies of such programs do not meet … quality standards.
So why does research not back it up completely? Is some piece — or pieces — perhaps missing from the OG approach?
2. Dyslexia treatment and cognitive training
Edublox’s dyslexia treatment program targets cognitive skills, a missing piece of the OG approach.
“Cognition” is defined as “the act or process of knowing.” Cognitive skills, therefore, refer to those skills that make it possible for us to know. They have more to do with the mechanisms of how we learn than with any actual knowledge.

A body of evidence has shown that children with dyslexia struggle not only in reading, spelling, writing, and reading comprehension but have multiple cognitive deficits. Cognitive psychology has now linked many brain-based skills to dyslexia: phonological awareness; verbal fluency; attention and executive functions; visual attention; visuospatial abilities; processing speed; short-term memory; auditory working memory; visual memory and visual sequential memory; visual long-term memory, especially for details; verbal long-term memory; and rapid naming.
Weak cognitive skills prevent a process called orthographic mapping. Every word has three forms: its sounds, spelling, and meaning. Orthographic mapping involves the brain linking the three word forms and storing them together in long-term memory. Orthographic mapping allows for instant word recognition, fluent reading, and accurate spelling.
Unfortunately, cognitive training, popularly known as brain training, has fallen into disfavor among learning disability practitioners. In one study that contributed to the distrust of brain training, viewers of the BBC science program Bang Goes the Theory were recruited to practice a series of online tasks for a minimum of ten minutes a day, three times a week, for six weeks.
A total of 11,430 volunteers aged 18 to 60 completed the study. Although they improved on the tasks, “no evidence was found for transfer effects to untrained tasks, even when those tasks were cognitively closely related” (Owen et al., 2010).
One reason why the experiment yielded no transfer effects is simply that the training time was too short. Subjects completed an average of 24 sessions — at ten minutes a session, that is, only four hours of training. According to fitness authority Bobby Maximus (2018), getting physically fit requires at least 130 hours. The brain is no different; even ten hours would have yielded little to no results.
3. Dyslexia treatment targeting two brain regions
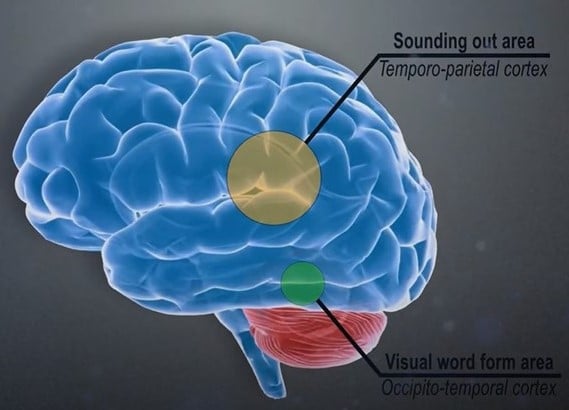
Research shows that a network of brain regions is involved in learning to read, one specifically in sounding out words and another in seeing words as pictures. The left inferior parietal lobule is involved in word analysis, grapheme-to-phoneme conversion, and general phonological and semantic processing. The picture area is located in the left occipitotemporal region and is known as the visual word form area (VWFA) or visual dictionary.
In the largest study of its kind up to date, Brem et al. (2020) confirmed that the VWFA is key to fluent reading. In addition, research by Pedago et al. (2010) supports that the VWFA plays a crucial role in the discrimination of letter orientation, which links to the well-known fact that a deficit in discriminating between a b and a d is one of the common symptoms of dyslexia.
Given that the most robust underactivation in children with dyslexia occurs in the occipitotemporal cortex, this brain region should be a prime target for treating dyslexia. Yet, so far, the VWFA has been neglected in dyslexia treatment.
Edublox’s dyslexia treatment program is aligned with the Orton-Gillingham approach, which is excellent at developing the sounding out area. However, our program simultaneously targets the brain’s VWFA, which is crucial for fluent reading.
4. Dyslexia treatment and learning principles

The application of certain educational learning principles is a core aspect that has so far been overlooked in dyslexia interventions. Thorndike (1874–1949) called learning principles the “laws of learning” and provided a conceptual foundation for how people learn. Examples of Edublox’s “laws of learning” in dyslexia treatment are listed below. Applying these fundamental learning principles contributes to helping children with dyslexia learn to read and write.
4.1) Learning is a stratified process
This age-old — but ageless — principle was already pointed out by Herbart (1776–1841). “One never, he insisted, apprehends anything in isolation, but always in terms of one’s background of previous experience and learning. So the first consideration in properly organized learning would be to ensure that the learner had the right background” (Mursell, 1954, pp. 210-211). Putting it differently, students’ prior skills and knowledge can help or hinder learning. Specific preliminary skills need to be mastered first, before anyone can learn the skills of adding and subtracting or the game of baseball. The same applies to reading.
4.2) Cognitive skills training should be multi-cognitive
In physical training, a balanced workout is vital as overtraining one part of the body can cause deformity, for example, the Popeye syndrome when overtraining the biceps. The brain is no different. For example, in Maguire et al.’s experiment with London taxi drivers, growth in the posterior hippocampi seems to have come at a cost, as they had reduced anterior hippocampal gray matter volume compared with bus drivers, with anterior volume decreasing with more navigation experience (Maguire et al., 2006).
One should also consider the role of mutualism. A mutualistic view suggests that cognitive abilities mutually facilitate growth. For example, better reasoning skills allow individuals to improve their vocabulary more quickly, and better vocabulary is associated with faster improvement in reasoning ability (Kievit et al., 2017).
4.3) Cognitive skills can only be improved by task loading
In the classic experiment of Ericsson and Chase, transfer did not occur following 250 hours of short-term memory training. The student, known as SF, was required to repeat sequences of random digits. Every time he got a sequence right, a digit was added. When the experiment was concluded, SF could repeat sequences of a staggering eighty-plus random digits. However, when tested on random alphabet letters, his short-term memory performance immediately reverted to normal.
Through interviews, the researchers discovered that SF, a competitive runner, pictured an unconnected string of three or four digits as one single race time. In this way, he relieved the burden on short-term memory and, like any mnemonist, took advantage of an already existing code in long-term memory (Shenk, 2010).
One way to prevent students from using techniques or strategies is to offer a series of cognitive exercises.
4.4) New knowledge and skills cannot be learned without repetition
As far as one can go back in history, repetition (also known as drill-and-practice) has been considered the backbone of successful teaching and schooling. However, in the 1920s and 1930s, repetition and rote learning came to define bad teaching. Good teachers do not fire off quiz questions and catechize kids about facts. They do not drill students on spelling or arithmetic.
When properly conducted, however, drill-and-practice is a consistently effective teaching method and should not be slighted as “low level,” and appears to be just as essential to complex and creative intellectual performance as they are to the performance of a virtuoso violinist (Brophy, 1986).
Whatever the case for the average student, repetition certainly benefits the child with learning challenges. For example, a meta-analysis of 85 academic intervention studies with learning-disabled students found that regardless of the practical or theoretical orientation of the study, the largest effect sizes were obtained by interventions that included systematic drill, repetition, practice, and review (Swanson & Sachse-Lee, 2000).
From a neuroscientific perspective, repetition is essential in the “wiring” of a person’s brain, i.e., forming connections or synapses between the brain cells. Without repetition, key synapses do not form. Furthermore, if such connections, once formed, are used too seldom to be strengthened and reinforced, the brain, figuring they are dead weight, eventually “prunes” them away (Hammond, 2015; Bernard, 2010).
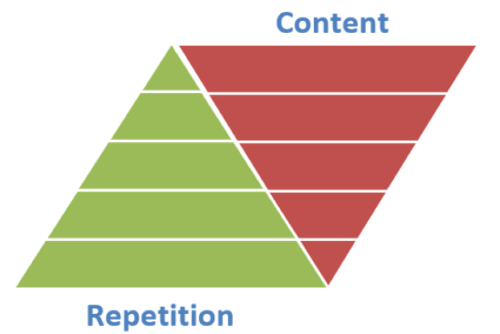
4.5) A “pyramid of repetition” needs to be constructed for the beginner learner
This principle, derived from the works of Suzuki, means that the beginner learner must start by repeating a limited amount of material many times over and over. Gradually, less and less repetition will be necessary to master new skills and knowledge.
In his book Nurtured by Love, Suzuki (1993) shares the story of a little parakeet taught to say in Japanese: “I am Peeko Miyazawa, I am Peeko Miyazawa.” According to Mr. Miyazawa, Peeko’s trainer, it took three thousand repetitions of the word Peeko over two months before the bird began to say “Peeko.” After that, “Miyazawa” was added, and after having heard “Peeko Miyazawa” only two hundred times, the little bird could say its full name. Then, when Mr. Miyazawa coughed for some days due to a cold, Peeko would say his usual “I am Peeko Miyazawa” and then cough. With less and less repetition, Peeko learned more and more words and also learned to sing.
“Once a shoot comes out into the open, it grows faster and faster… talent develops talent and… the planted seed of ability grows with ever-increasing speed” (Suzuki, 1993, p. 6).
No doubt it is the same with a human being, as the following story, also taken from Suzuki (1993, pp. 92-93), illustrates:
Since 1949, our Mrs. Yano has been working with new educational methods for developing ability, and every day she trains the infants of the school to memorize and recite Issa’s well-known haiku. [A haiku is a short Japanese poem consisting only of three lines.]… Children who at first could not memorize one haiku after hearing it ten times were able to do so in the second term after three to four hearings, and in the third term after only one hearing.
The bottom line
By combining cognitive training with the systematic teaching of reading that targets both the brain’s sounding out area and the VWFA, while applying fundamental learning principles, the way is paved to improve the reading and writing abilities of children with dyslexia — and help them overcome their challenges.
Watch our playlist of customer reviews and experience how Edublox’s dyslexia treatment program helps overcome dyslexia signs and symptoms, and book a free consultation to discuss your child’s learning needs. We cater to a variety of dyslexia types.
Key takeaways
Authored by Susan du Plessis (B.A. Hons Psychology; B.D.), a reading specialist with 30+ years of experience in the learning disabilities field.
Medically reviewed by Dr. Zelda Strydom (MBChB).