Table of contents:
- Introduction
- Neuroplasticity: An extraordinary discovery
- Brain differences in the dyslexia brain
- Poor connectivity in the dyslexia brain
- Reduced plasticity in the dyslexia brain
- Do brain differences equal brain disorders?
- Conclusion
.
Introduction
Developmental dyslexia is a neurodevelopmental learning disorder that affects an estimated 5 to 20% of the U.S. population. Its defining feature is difficulty with decoding and recognizing words—challenges that can’t be explained by lack of instruction, poor motivation, or low intelligence.
The concept that dyslexia is related to the brain dates back to the late 1800s. Researchers such as Berlin (Opp, 1994), Morgan (1896), and Hinshelwood (1895) suggested that reading difficulties might stem from brain dysfunction or injury. These early theories gained traction in the 1980s and 90s when autopsy studies seemed to support the idea.
One influential study by Galaburda and colleagues (1985) reported brain abnormalities in a small group of people with a history of dyslexia. But the study had serious limitations. The sample was tiny—only seven people—and included individuals with other neurological or psychiatric conditions. Some had broader cognitive impairments that extended beyond reading. The control group was also very small, making it hard to know whether the findings were truly unique to dyslexia.
At the time, the dominant belief in neuroscience was that the brain was hardwired. If someone was born with a learning difficulty or suffered brain damage, it was widely assumed that little could be done to change the situation (Keshav, 2018). This mindset helped reinforce the idea — still common today — that dyslexia is a permanent condition. But as later discoveries would show, the brain is far more adaptable than anyone imagined.
Neuroplasticity: An extraordinary discovery
A landmark 1998 study found that the human brain can generate new neurons (Eriksson et al., 1998). This discovery challenged the long-held belief that the brain is a rigid system — that humans are born with all the brain cells they will ever have and that lost cells are never replaced. This groundbreaking discovery sparked a wave of research demonstrating that the brain is not rigid but rather adaptable. It can reorganize, grow, and even compensate for damage — a property known as neuroplasticity.

One of the most compelling demonstrations of this plasticity came from a study by Maguire et al. (2000), who investigated the brains of London taxi drivers. The idea originated from animal research, where scientists observed that the hippocampus — a brain structure crucial for memory and spatial navigation — was larger in animals that stored food for later use. In some species, the hippocampus even grew during times when memory demands were highest.
Maguire wondered whether the same might be true in humans who relied heavily on memory for navigation. London cab drivers, after all, must memorize an estimated 25,000 streets and thousands of landmarks to earn their licenses. When Maguire compared the brains of cab drivers with those of non-taxi drivers, the results were striking: the cab drivers had significantly larger posterior hippocampi. In a follow-up study, she compared them to London bus drivers, who drive fixed routes, and found the same pattern. The more years of navigation experience a taxi driver had, the larger their hippocampal gray matter volume (Maguire et al., 2006).
Similar findings have been observed in other groups where intensive learning or practice is involved. Musicians (Sluming et al., 2002), jugglers (Draganski et al., 2004), and bilinguals (Mechelli et al., 2004) all show structural brain differences that appear to reflect their specialized training. In each case, the more time spent practicing, the more pronounced the brain changes become.
Together, these studies show that the brain is not only capable of changing in response to learning and experience but also of reorganizing itself in the face of injury. Research by Nudo (2013) and Kadosh and Walsh (2006) has shown that healthy brain areas can sometimes take over the roles of damaged regions—a powerful reminder of the brain’s potential to adapt and heal.
Brain differences in the dyslexia brain
As technology advanced, neuroscience began to play a larger role in dyslexia research. Unfortunately, many brain imaging studies in this area still suffer from small sample sizes and inconsistent findings, making it difficult to draw firm conclusions (Protopapas & Parrila, 2018). However, some patterns have emerged. In a meta-analysis of functional neuroimaging studies, Martin et al. (2016) identified several brain regions that consistently differed between people with and without dyslexia. The most reliable differences were observed in the left occipitotemporal cortex, home to a region known as the visual word form area (VWFA), believed to be critical for fluent reading.
Researchers at Georgetown University Medical Center investigated how this region works in skilled readers. They found that the VWFA (highlighted in yellow in brain images) stores written words in a sort of mental “visual dictionary,” allowing readers to recognize them almost instantly (Glezer et al., 2016). Using fMRI scans, Glezer and colleagues studied how the brain responds to words that sound the same but look different — such as hare and hair. They discovered that these words activated entirely different sets of neurons in the VWFA, just like separate entries in a visual catalog. If sound had influenced this region, similar-sounding words would have triggered similar activity. However, hare and hair looked just as different in the brain as hair and soup.
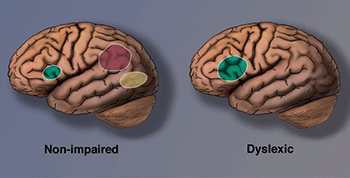
At the same time, the researchers identified a separate brain region sensitive to sound — one where hare and hair did look the same. These results highlight the brain’s division of labor during reading: one region handles the visual aspects of words, while another processes the phonological (sound-based) side.
In the most extensive neuroimaging study on dyslexia to date, Brem et al. (2020) confirmed that the VWFA plays a key role in fluent reading. Their study examined fMRI data from 140 children (ages 7.9 to 12.2), including 55 children with dyslexia, 73 typical readers, and 12 intermediate readers. The results showed that the connectivity of the left occipitotemporal cortex is essential for the VWFA to specialize effectively. Supporting this, Pegado et al. (2011) found that the VWFA is also involved in distinguishing between similar-looking letters, such as b and d — a skill often impaired in dyslexia.
Given that underactivation in the left occipitotemporal cortex is one of the most consistent findings in children with dyslexia (Brem et al., 2020; Martin et al., 2019), this brain region is a promising target for interventions to improve reading fluency.
These findings challenge the dual-access theory (Frost, 1998), which proposes that skilled readers rely on both phonology and visual processing when recognizing words. Instead, the evidence suggests that the brain has distinct systems for each function — and that the VWFA operates independently of sound processing.
Another key region identified by Martin and colleagues is the left inferior parietal lobule (shown in red), which is involved in phonological decoding, grapheme-to-phoneme conversion, and semantic processing.
Meanwhile, brain scans also reveal signs of compensatory overactivation in other regions of the brain’s reading network (highlighted in green). These areas help people with dyslexia read more accurately. However, since the visual word form area remains underactive, difficulties with fast, fluent, automatic reading tend to persist. As Shaywitz (2005) notes, many individuals with dyslexia can become accurate readers, but they often continue to read slowly.
Reduced plasticity in the dyslexia brain
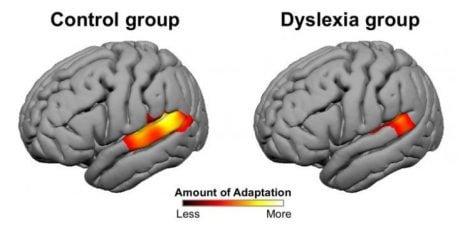
While neuroplasticity is one of the brain’s great strengths, some researchers have found that this capacity for change may function differently in individuals with dyslexia. A study by Perrachione and colleagues (2016) at MIT revealed that neural adaptation — the brain’s ability to adjust its response to repeated stimuli — is significantly reduced in people with dyslexia.
Using MRI, the researchers observed how young adults with and without dyslexia responded to repeated spoken words. In typical readers, certain language-related brain areas showed a tuning effect: when the same speaker repeated a word, these areas became less active, indicating adaptation. However, when the speaker changed, the brain responded afresh — as expected. In contrast, individuals with dyslexia showed far less adaptation, even when the voice and word remained the same.
In a second task, participants viewed repeated or varying words, objects, and faces. Again, the brains of those with dyslexia showed reduced adaptation — not only in regions related to language but also in areas involved in general visual and object recognition.
These findings suggest that the difference is not just about reading or spoken language. Instead, it may reflect a broader difficulty in perceptual learning — the brain’s ability to fine-tune itself through exposure and repetition.
Perhaps most importantly, this reduced adaptability has also been observed in young children. When researchers tested first- and second-graders with dyslexia, the same diminished neural adaptation was present, indicating that this trait may be part of the developmental foundation of dyslexia rather than a consequence of reading failure.
Poor connectivity in the dyslexia brain
A groundbreaking study from the MRC Cognition and Brain Sciences Unit at the University of Cambridge suggests that learning difficulties, such as dyslexia, may not stem from isolated brain regions. Instead, the problem could lie in how different parts of the brain are connected (Siugzdaite et al., 2020).
To explore this idea, researchers used machine learning to analyze the brain scans of 479 children. Of these, 337 had been referred for learning-related cognitive challenges, and 142 formed a comparison group. The team combined neuroimaging data with a wide array of cognitive, learning, and behavioral assessments.
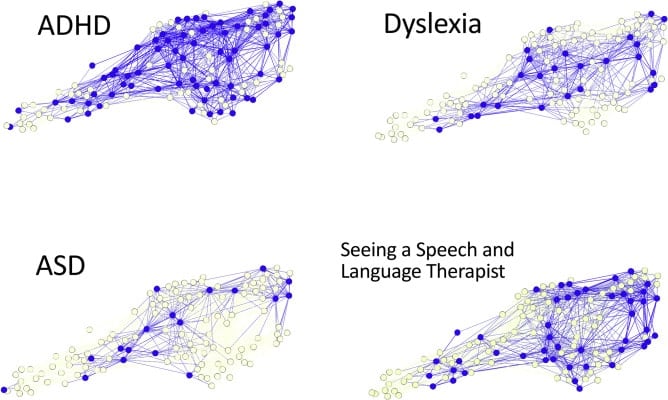
What they found was surprising: the brain differences they identified did not align with diagnostic labels. In other words, no specific brain region reliably predicted whether a child had dyslexia, ADHD, or autism spectrum disorder. Even more striking, there were no consistent neural patterns associated with specific cognitive impairments, such as language or memory.
Instead, the researchers discovered that each child’s brain was organized around networks of hubs — much like the key junctions in a transportation system. Children with well-connected hubs tended to have either no learning problems or very specific, isolated difficulties (like trouble with listening). In contrast, children whose brain hubs were poorly connected — like train stations with limited routes — often had broader, more severe learning challenges across multiple domains.
This study shifts the focus away from searching for a single “dyslexia spot” in the brain. Instead, it supports a systems-level view: learning difficulties may arise not from broken parts but from weakened connections between them.
Do brain differences equal brain disorders?
While Protopapas and Parrila (2019) emphasize that poor reading is a serious issue that requires early assessment and intervention, they caution against assuming that differences in the brain automatically indicate a disorder. In fact, after reviewing decades of scientific research, they came to a very different conclusion.
In their 2018 review, they write:
Differences in brains are certain to exist whenever differences in behavior exist, including differences in ability and performance. Therefore, findings of brain differences do not constitute evidence for abnormality; rather, they simply document the neural substrate of the behavioral differences. We suggest that dyslexia is best viewed as one of many expressions of ordinary ubiquitous individual differences in normal developmental outcomes. Thus, terms such as “dysfunctional” or “abnormal” are not justified when referring to the brains of persons with dyslexia.
In other words, just because brain scans show differences between dyslexic and typical readers does not mean those differences are inherently faulty or pathological. They may simply reflect natural variation in how people develop — especially when it comes to learning.

What is more, some brain differences may not be causes of dyslexia at all but rather consequences of struggling to read. In a study published in the Journal of Neuroscience, Krafnick et al. (2014) compared three groups of children: one group with dyslexia, an age-matched group without dyslexia, and a younger group matched for reading level. While children with dyslexia showed less gray matter than typical readers of their age, they had similar gray matter volumes to those of younger, lower-level readers. This suggests that some of the anatomical differences observed in dyslexia may result from limited reading experience rather than an underlying brain defect.
Additional support for this idea comes from Huettig et al. (2018), who note that many of the deficits linked to dyslexia — such as problems with phonological awareness or verbal memory — are also common in illiterate or minimally literate adults. These deficits, they argue, often stem from a lack of exposure to reading rather than a fixed cognitive flaw.
In fact, a study by Skeide et al. (2016) showed that when illiterate adults learned to read for the first time, the changes in their brains extended far beyond the cortex. After just six months of literacy instruction, the participants — Indian women with no prior schooling — showed structural changes deep in the brain, including in the thalamus and brainstem. This underscores just how profoundly reading experience can shape the brain.
Taken together, these findings challenge the idea that brain differences automatically signal something abnormal. They remind us that the brain is shaped by what we do — and what we learn — as much as by what we’re born with.
Conclusion
There are indeed brain differences between people with dyslexia and typical readers. But whether those differences are the cause of reading struggles — or a result of them — is still not conclusive. What we do know, and what gives immense hope, is that the brain is plastic. It can change. It can adapt. It can grow.
The human brain is a powerhouse of possibilities. It has launched astronauts to the moon, engineered microchips that perform billions of calculations per second, and invented a simple system of red, yellow, and green lights to guide millions of people through traffic every day. Most astonishing of all, it has even learned how to examine itself — to peer into its own circuitry and discover how to rewire when something isn’t working right.
So while scientists continue to explore the roots of dyslexia and its neural signature, one truth remains clear: the brain tells us — through its very structure and function — that it is capable of overcoming obstacles, including those linked to dyslexia. With the right interventions, consistent practice, and an environment that believes in change, struggling readers can build the neural pathways they need to thrive.
Dyslexia may begin in the brain, but it doesn’t end there. It ends — or rather, it transforms — through effort, support, and the extraordinary capacity of the human mind to learn, adapt, and grow.
Edublox offers explicit, systematic reading instruction integrated with cognitive training to help students overcome the symptoms of dyslexia. We work with families worldwide. Book a free consultation to explore how we can support your child’s learning journey.
References for Dyslexia and the Brain: What Science Reveals:
- Brem, S., Maurer, U., Kronbichler, M. et al. (2020). Visual word form processing deficits driven by severity of reading impairments in children with developmental dyslexia. Scientific Reports, 10.
- Draganski, B., Gaser, C., Busch, V., Schuierer, G., Bogdahn, U., & May, A. (2004). Neuroplasticity: Changes in grey matter induced by training. Nature, 427, 311-2.
- Eriksson, P. S., Perfilieva, E., Björk-Eriksson, T., Alborn, A., Nordborg, C., Peterson, D. A., Gage, F. H. (1998). Neurogenesis in the adult human hippocampus. Natural Medicine, 4(11), 1313-7.
- Frost, R. (1998). Toward a strong phonological theory of visual word recognition: True issues and false trails. Psychological Bulletin, 123(1), 71-99.
- Galaburda, A. M., Sherman, G. F., Rosen, G. D., Aboitiz, F., & Geschwind N. (1985). Developmental dyslexia: Four consecutive patients with cortical anomalies. Annals of Neurololgy, 18(2), 222-33.
- Glezer, L. S., Eden, G., Jiang, X., Luetje, M., Napoliello, E., Kim, J., & Riesenhuber, M. (2016). Uncovering phonological and orthographic selectivity across the reading network using fMRI-RA. Neuroimage, 138, 248-56.
- Hinshelwood, J. (1895). Word-blindness and visual memory. Lancet, 146(3773), 1564-70.
- Huettig, F., Lachmann, T., Reis, A., & Petersson, K. M. (2018). Distinguishing cause from effect—Many deficits associated with developmental dyslexia may be a consequence of reduced and suboptimal reading experience. Language, Cognition and Neuroscience, 33(3), 333–50.
- Kadosh, R. C., & Walsh, V. (2006). Cognitive neuroscience: Rewired or crosswired brains? Current Biology, 16(22), R962-3.
- Keshav, M. (2018). The life transforming power of NLP. Chennai: Notion Press.
- Krafnick, A. J., Flowers, D. L., Luetje, M. M., Napoliello, E. M., & Eden, G. F. (2014). An investigation into the origin of anatomical differences in dyslexia. Journal of Neuroscience, 34(3).
- Maguire, E. A., Gadian, D. G., Johnsrude, I. S., Good, C. D., Ashburner, J., Frackowiak, R. S. J., & Frith, C. D. (2000). Navigation-related structural change in the hippocampi of taxi drivers. Proceedings of the National Academy of Sciences of the United States of America, 97(8), 4398-403.
- Maguire, E. A., Woollett, K., & Spiers, H. J. (2006). London taxi drivers and bus drivers: A structural MRI and neuropsychological analysis London taxi drivers and bus drivers. Hippocampus, 16(12), 1091-101.
- Martin, A., Kronbichler, M., & Richlan, F. (2016). Dyslexic brain activation abnormalities in deep and shallow orthographies: A meta-analysis of 28 functional neuroimaging studies. Human Brain Mapping, 37, 2676–99.
- Martin, L., Durisko, C., Moore, M. W., Coutanche, M. N., Chen, D., & Fiez, J. A. (2019). The VWFA is the home of orthographic learning when houses are used as letters. eNeuro, 6(1).
- Mechelli, A., Crinion, J. T., Noppeney, U., O’Doherty, J., Asburner, J., Frackowiak, R. S., & Price, C. J. (2004). Structural plasticity in the bilingual brain. Nature, 431, 757.
- Morgan, W. P. (1896). A case of congenital word blindness. British Medical Journal, 2(1871), 1378.
- Nudo, R. J. (2013). Recovery after brain injury: Mechanisms and principles. Frontiers in Human Neuroscience, 7(887).
- Opp, G. (1994). Historical roots of the field of learning disabilities: Historical roots of the field of learning disabilities: Some nineteenth-century German contributions. Journal of Learning Disabilities, 27, 10-9.
- Pegado, F., Nakamura, K., Cohen, L., & Dehaene, S. (2011). Breaking the symmetry: Mirror discrimination for single letters but not for pictures in the Visual Word Form Area. Neuroimage, 55, 742-9.
- Perrachione, T. K. , Del Tufo, S. N., Winter, R., Murtagh, J., Cyr, A., Chang, P., … Gabrieli, J. D. E. (2016). Dysfunction of rapid neural adaptation in dyslexia. Neuron, 92(6), 1383-97.
- Protopapas, A., & Parrila, R. (2018). Is dyslexia a brain disorder? Brain Sciences, 8(4).
- Protopapas, A., & Parrila, R. (2019). Dyslexia: Still not a neurodevelopmental disorder. Brain Sciences, 9(1).
- Shaywitz, S. (2005). Overcoming dyslexia. New York: Vintage Books.
- Siugzdaite, R., Bathelt, J., Holmes, J., & Astle, D. E. (2020). Transdiagnostic brain mapping in developmental disorders. Current Biology, 30(7).
- Skeide, M. A., Kumar, M., Mishra, R. K., Tripathi, V. N., Guleria, A., Singh, J.P., … & Huettig, F. (2017). Learning to read alters cortico-subcortical cross-talk in the visual system of illiterates. Science Advances, 3(5).
- Sluming, V., Barrick, T., Howard, M., Cezayirli, E., Mayes, A., & Roberts, N. (2002). Voxel-based morphometry reveals increased gray matter density in Broca’s area in male symphony orchestra musicians. Neuroimage, 17(3), 1613–22.
Dyslexia and the Brain: What Science Reveals was authored by Sue du Plessis (B.A. Hons Psychology; B.D.), an educational specialist with 30+ years’ experience in the field of learning disabilities. Medically reviewed by Dr. Zelda Strydom (MBChB).
Edublox is proud to be a member of the International Dyslexia Association (IDA), a leading organization dedicated to evidence-based research and advocacy for individuals with dyslexia and related learning difficulties.

